Fundamentos de cinética enzimática
Para poder modelar matemáticamente la velocidad de reacción (cinética) catalizada por enzimas, se toman ciertas suposiciones:
1. Solo hay un sustrato y solo hay un producto
Si consideramos la reacción de la enzima, con unión a sustrato y luego generación de productos:
Puede que haya más pasos intermedios, pero se considera que el más relevante es la unión de la enzima con el sustrato. Así que se hacen dos supuestos:2. Paso limitante es la unión de la enzima al sustrato, y por tanto, la formación del estado de transición es la limitante.
3. El lapso de tiempo elegido incluye todos los procesos y los engloba como uno solo.
Si hubiera pasos intermedios más rápidos que el de la formación del complejo Enzima-Sustrato (ES), dado que la cinética dependerá de la formación de este, y éste es el más "lento", podemos despreocuparnos de los que son más rápidos porque el modelo no dependerá de ellos, dado que ocurrirán en lapsos de tiempo menores. Esta parte es interesante, porque entonces si trabajas a velocidades lentas, es decir, tiempos relativamente grandes, muchos procesos biológicos de formación de complejos, pueden modelarse con estas mismas ecuaciones, por ejemplo, la unión de sustrato a microorganismo en un cultivo celular (de organismos unicelulares), o la adsorción de un sustrato/ligando sobre una resina y así.
De esta manera, nos centramos en las reacciones de disociación del complejo que es el limitante:
Para simplificar el modelo, estudiamos con el menor número de variables, así que si lo hacemos a tiempos donde apenas inicia la reacción, haremos que la concentración de producto sea tan pequeña o nula, así que el producto no interferirá con las determinaciones. De esta manera:
4. Cuando se determina la velocidad, no hay producto presente.Así, en la ecuación de arriba, no nos preocuparemos por P.
Con lo que:

......... (Ecuación 1)
Con los supuestos anteriores, entenderemos que la velocidad de la enzima depende de qué tanto de este complejo se genere, pues una vez generado, la liberación del producto ocurre tan rápido (o al menos más rápido que el paso limitante) que:
5. La velocidad de enzima unida es solo proporcional de la cantidad de enzima unida.
Si consideramos esto, llamemos fu a la fracción de enzima que está unida, fu=[ES]/[E]total:
Por lo tanto,
......... (Ecuación 2)
Recordemos que la concentración total de la enzima es igual a la suma de la concentración de enzima libre y la concentración de enzima unida (en el complejo ES), así que
y
por lo que si sustituimos esto en la Ecuación 1, nos queda:
Arreglando un poco la ecuación:

......... (Ecuación 3)
Nota que en la ecuación 3, tenemos del lado izquierdo el equivalente de la fracción unida. De esta manera, si sustituimos esta Ecuación3, en la ecuación 2, nos queda:

.........(Ecuación 4)
Nota que puse Km en lugar de K, esta es la ecuación de Michaelis-Menten.
En lugar de Vmax, también se puede utilizar actividad enzimática (que es la velocidad/mg de proteína)
La Ecuación 4, tiene forma hiperbólica, como ocurre con todas las ecuaciones de adsorción.
Para ajustarlo, se puede hacer con los programas computacionales que utilizan algoritmos matemáticas iterativos, Sin embargo, antes para calcular el valor de la Km y Vmax, se tenía que linearizar la ecuación, esto se logra invirtiendo ambos lados de la ecuación.
Rearreglando:
Eso significa que se puede ajustar a una línea,
donde
 y
y 
Por lo que
así que despejando:
y
Así que Vmax es la inversa de la ordenada al origen.
De esta manera, esta linearización que es llamada Lineweaver-Burk, te da datos que se pueden modelar por regresión lineal por mínimos cuadrados:
Nota que hay dos problemitas, por un lado, mientras más pequeño es el valor del sustrato, más se aleja y entonces tiene más peso a la hora de calcular la ordenada al origen. Por otro, como a menor valor del sustrato, la posibilidad de error es mayor, es probable que ese error afecte el resultado de la ordenada al origen y por tanto de la Vmax.
De ahí que es mejor utilizar programas computacionales con algoritmos iterativos para calcular sin tener que transformar los datos a línea.
Ahora bien, qué ocurriría si se uniera más de un sustrato por enzima, por ejemplo, que se una a n moléculas de sustrato, entonces la ecuación 1 se convertiría a
Por lo que al hacer el mismo proceso, la Ecuación 4 se convierte a:
Que es la llamada Ecuación de Hill:
Con esto, al modelar es posible saber si la enzima tiene más de un sitio de unión, pero a veces, aunque tenga, por ejemplo, 2 sitios, la h no da de 2 sino de 1.5, esto es porque hay cooperatividad positiva, es decir el segundo sitio, incrementa su afinidad, es decir su Km disminuye, por lo que entonces el segundo valor aparenta como si fuera Km^0.5. Como también puede ocurrir cooperatividad negativa, en números superiores sería un problema. Porque la cooperatividad negativa, haría que la h fuera mayor a la n, es decir, si tuviera 3 sitios disponibles, pero hay cooperatividad negativa, el segundo sitio tendría menos afinidad que el primero, por lo que su Km sería mayor, y lo mismo con el tercero, así que la h sería mayor a 3.
Por ejemplo, Imagina que te sale una h de 3.5, cómo sabes si viene de una cooperatividad positiva de 4, o de una cooperatividad negativa de 3. Bueno, para ello se ha propuesto un índice que es el índice R que toma en cuenta las concentraciones de Sustrato para alcanzar el 90% de la Vmax (S90 o S0.9) y para alcanzar el 10% de la Vmax (S10 o S0.1):
Estos dos valores son más fáciles de alcanzar experimentalmente
Cuando no hay más de un sitio, R=81, cuando hay cooperatividad positiva R<81 y cuando hay cooperatividad negativa R>81.
R se relaciona con h mediante la fórmula siguiente:

Sin embargo, si h >2 y la cooperatividad es negativa, la interpretación de R se complica un poco. La ventaja de R, es que no requiere del modelado, sino que se puede calcular a partir de los datos experimentales directos.
Eficiencia catalítica
Para poder tener una idea de la velocidad de la enzima a nivel molecular, se puede saber a partir de cuántas moles de sustrato cataliza por moles de enzima, en lugar de hacerlo sobre gramos o miligramos de enzima. A este concepto se le llama número de
recambio enzimático kcat.

Esto nos da idea de la cantidad en moles de sustrato que cada molécula de enzima puede catalizar y por tanto cuántos sustratos pueden interactuar por molécula de enzima. Así que las unidades de kcat son en t-1 por ejemplo s^-1.
No obstante, para poder calcularlo se requiere que la proteína esté pura o lo más puro posible para que la cantidad de enzima pueda cuantificarse lo más exacto sin subestimarlo por la presencia de otras proteínas.
La ecuación de Vmax, quedaría:

........ (Ecuación 5)
Este valor nos permite poder comparar enzimas aún de diferente sustrato, pues nos permite hacer dicha comparación sobre la base de cuántos sustratos puede catalizar cada molécula de enzima. De ahí que se puede observar qué tan eficiente es una enzima si su kcat es lo suficientemente grande para no depender de su equilibrio de disociación Km.
Por lo que se toma la relación siguiente para evaluar la eficienca catalítica o perfección catalítica. Esta relaciona la velocidad de catálisis por molécula de enzima, entre la constante de disociación, es decir, que es afectada por las tendencias al equilibrio, así como por la velocidad de difusión del sustrato en la solución.
Nota que las unidades de la eficiencia catalítica son en M^-1 s^-1. Una interpretación es qué tantos moles de sustrato puede catalizar cada molécula de sustrato relativo a las veces en que se puede disociar, o cuántas veces las moles de producto generadas respecto a la disociación de productos. También puede interpretarse como qué tan rápida es la generación del producto (segundo paso de la reacción 1) respecto al total del proceso catalítico.
Km y Vmax, nos permite comparar enzimas que catalizan la misma reacción. Mientras que kcat y Eficienca catalítica nos permite evaluar a enzimas aún si son diferentes sus reacciones.
Comparemos diferentes enzimas y veamos su eficienca catalítca
Eficiencia catalítica (10^6 M^-1 s^-1)
Acetilcolinesterasa 140
Anhidrasa carbónica 15
Catalasa 0.09
Cetosteroide isomerasa 300
Fumarasa 160
β-lactamasa 100
Orotidina-monofosfato descarboxilasa 56
Pepsina 0.001 7
Ribonucleasa 0.1
Quimotripsina 0.000 0093
Triosa fosfato isomerasa 240
tRNA sintetasa 0.008 4
La Anhidrasa carbónica es de las más eficientes, cataliza la generación de productos más de 15,000,000 veces más rápido que la velocidad con la que el sustrato se asocia. Podríamos decir, que, en cuanto el sustrato se asocia, casi "instantáneamente" se genera el producto. Así que en efecto, el paso limitante o más lento, es la unión con el sustrato.
Generalmente, el límite superior de la eficiencia catalítica es mayor a 100.10^6 M^-1 s^-1
Por ejemplo observemos la gráfica de Shafee: Par Thomas Shafee — Travail personnel, CC BY 4.0, https://commons.wikimedia.org/w/index.php?curid=29353835, donde graficó las eficiencias catalíticas enzimáticas respecto a la cantidad de enzimas observadas en ese grupo
Puedes ver que pocas enzimas son clasificadas como perfectas catalíticamente, Son las encerradas en el rectángulo gris.
¿Qué pasaría si hubiera más estados de Transición?
Buenos se podría modelar, si suponemos que el cambio conformacional, hace que en esos tiempos, la reacción del ajuste inducido no sea reversible, y entonces : Este es el modelo de
Briggs-Haldane. La constante de Km cambiaría a:
y la Ecuación 5 devendría a:
Con lo que kcat sería:
Actualmente, ya las ecuaciones en general no se resuelven así, sino sobre su definición cinética pero como son varios pasos, ya se trabaja de forma matricial. Por ejemplo:
para la reacción:
Pero eso será materia de otro blog...


 ......... (Ecuación 1)
......... (Ecuación 1)
 ......... (Ecuación 2)
......... (Ecuación 2)





 ......... (Ecuación 3)
......... (Ecuación 3) .........(Ecuación 4)
.........(Ecuación 4)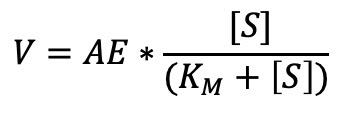





 y
y 











 ........ (Ecuación 5)
........ (Ecuación 5)






No hay comentarios:
Publicar un comentario